
|
|
|
Главная -> Потоки электромагнитных излучений 0 [1] 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 логических тканей целого организма. Тканевые системы условно назовем биолосическими микросистемами, отдельные части тела (голову, туловище и т. д.) и организм в целом - биологическими макросистемами. 1.1. ВЗАИМОДЕЙСТВИЕ ЭМИ С БИОЛОГИЧЕСКИМИ МИКРОСИСТЕМАМИ По мере удаления от источника излучения свойства ЭМ-поля меняются. На расстоянии г , превьпиающем размеры источника d и длину вошы Х, а точнее - удовлетворяющем одновременно условиям г > Х/2я и г> > d/\ поле существует в виде сформировавщихся бегущих волн. При частоте колебаний ниже 3 10* Гц (300 МГц) в качестве характеристики поля принимают электрическую (Е) и магнитную (Н) составляющие при частоте колебаний вьпие 300 МГц ППЭ (/): / = iVl207r= I201rЯ Такой выбор характеристик исторически установился в радиотехнике в связи с особенностями методов измерения Е и Н на разных частотах. На меньщих расстояниях (в зоне индукции) может преобладать энергия электрического или магнитного полей. В этом случае Е и Н изменяются не в фазе и быстро убывают с увеличением расстояния от источника (1/г и 1/г соответственно), причем плотность энергии ЭМ-поля W (зрг/см) равна сумме плотностей электрического и магнитного Wff полей л-» £•• W„ = 210"*Я Кроме того, следует отметить характерные особенности искусственных излучателей - это высокая когерентность, т. е. частотная и фазовая стабильность, означающая значительную концентрацию энергии в узких областях спектра, и поляризация (например, радиовещательные передачи на ДВ, СВ и КВ ведутся с помощью вертикально поляризованных волн, а на ТВ, РВ и УКВ с ЧМ - горизонтально поляризованных волн). Поэтому изучение частотной зависимости и влияния поляризации на процессы поглощения ЭМ-знергии биологическими объектами представляется весьма актуальным. В отличие от ионизирующего излучения, непосредственно создающего электрические зарящ>и ЭМ-поля не обладают ионизирующей способностью и воздействуют только на уже имеющиеся свободные зяды или диполи. Диэлектрические свойства биологических тканей сильно зависят от их химического состава, частоты колебаний и интенсивности электрических процессов, происходящих внутри биологического объекта. ЭМч:войства определяют процессы прохождения энергии через слои вещества, отраженной на границах их раздела, и поглощения внутри тканей. Диэлектрические свойства мыщечной, кожной и других тканей с высоким содержанием воды, а также жировой, костной и других тканей с низким содержанием воды наиболее полно изучены Шваном и сотр. [76, 140, 141], Map- тином [121] и другими исследовагелями. Имеются обстоятельные обзоры по данной проблеме [33, 42, 80, 150]. При взаимодействии ЭМИ с биологическим веществом возникают два типа эффектов, определяющих диэлектрические свойства тканей. Колебания свободных зарядов (ионов) приводят к увеличению токов проводимости и потере энергии, связанной с электрическим сопротивлением среды. Вращение дипольных молекул с частотой приложения ЭМ-поля влияет на токи смещения и диэлектрические потери, обусловленные вязкостью среды. Соотношения между этими видами преобразования энергии выражаются комплексной диэлектрической проницаемостью или тангенсом угла потерь. Диэлектрические свойства биологических тканей достаточно полно можно описать относительной диэлектрической проницаемостью е и проводимостью а. Часто используемые для описания распространения ЭМ-волн в диэлектрике характеристики, такие как комплексная диэлектрическая проницаемость е* = б, -/ej и тангенс угла потерь tg5, можно выразить через б и а: е,=е; е2=а/(соео); tg5 = е/е, = o/(cjeeo), где со = 2я/ - круговая частота ЭМ-поля; Cq - электрическая постоянная, равная 8,85-10"Ф/м. Магнитные свойства тканей также можно охарактеризовать двумя величинами: относительной магнитной проницаемостью ц и магнитной проводимостью g (Вб/м), причем магнитная постоянная вакуума Цо-= 1,27-10"* Гн/м. Следует отметить, что хотя все биологические ткани с точки зрения их макроскопических свойств являются очень слабыми диа- и парамагнетиками, близкими по свойствам к воздушной среде (д s= 1 и g = 0), диссипация магнитной энергии в биологических объектах может быть значительной в зависимости от размеров и электрических свойств этих объектов. Например, при воздействии высокочастотного (10-15 МГц) магнитного поля в токопроводящих тканях организуются вихревые токи.-При этом вьщеляется количество тепла Q [79] Q = k где к - коэффициент, учитывающий размерности величины. Такое соотношение имеет максимум при а=/е/2, следовательно, в определенном диапазоне (3-300 МГц) можно подбирать частоты, при которьк нагревание тканей магнитным полем будет максимальным. Несмотря на то что в условиях целостного организма реализация этого явления в полной мере невозможна, его удается наблюдать при значительно различающихся частотах [76]. Диэлектрические свойства тканей существенно зависят от частоты ЭМ-колебаний (рис. 1.1). Зависимость диэлектрической проницаемости биологической ткани от частоты действующего поля условно можно разделить на три основные релаксационные области: а-дисперсия (Гц-кГц), -дисперсия (МГц) и у-дисперсия (ГГц). Такое деление обусловлено структурной сложностью живой материи. Раствор электролита проявляет 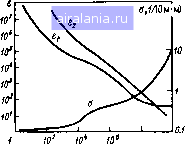 Рис. 1.1. Относ1Гтельиая диэлектрическая про-Яется ницаемость 6 и проводимость О биологических тканей с высоким содержанием вода в зависимости от частоты / ЭМ-колебаний е, и €2 - мнимые части комплексной диэлектрической проницаемости (140, 141] «V, Ги, ЛИШЬ 7-дисперсионную характеристику воды. Сложные, крупные субклеточные структуры обеспечивают -дисперсию, а релаксация зарядов у клеточньк мембран ответственна за «-релаксационную область. При воздействии на жань постоянного злектрического поля она поляризуется, и ионные токи будут протекать только по межклеточной жидкости, так как при постоянном напряжении мембраны клеток, являясь хорошими изоляторами, надежно изолируют внутриклеточное содержание. Под действием приложенного поля возможен электрофорез заряжен-ньк частиц, таких как макромолекулы и клетки. На частотах примернб меньше 10* Гц период ЭМ-колебаний достаточно большой, для того чтобы клеточные мембраны успели перезарядиться за счет ионов вне и внутри клетки. Это объясняет наличие низкой удельной ионной проводимости даже для тканей с высоким содержанием воды. При этом полный заряд и диэлектрическая проницаемость ткани за период колебаний также весьма велики. Последующий рост удельной проводимости происходит вследствие уменьшения емкостного сопротивления мембран с увеличением частоты. Неполная перезарядка изолированных мембран вовлекает внутриклеточную жидкость в процесс образования ионных токов, проводимость ткани плавно увеличивается, а ее дюлек-трическая проницаемость падает. Таким образом, релаксация противо-ионов у заряженных клеточных поверхностей является основным механизмом а-дисперсии [31, 76, 141]. Лавинное вовлечение внутриклеточной среды в процесс образования ионных токов на частотах Ю"* - 10* Гц вызывает резкое возрастание удельной проводимости. Кроме того, поляризация молекул тканей, в основном молекул воды, приводит к возникновению токов смещения, значительно увеличивающих токи в тканях при тех же амплитудах напряженности электрического поля, т. е. уменьшает их удельное сопротивление. /3-Дисперсию обусловливает неоднородность клеточной структуры биологической ткани. Поскольку такие образования, как митохондрии и ядро, малы по сравнению с самой клеткой, их релаксационные потери меньше, чем для клетки в целом. На частотах 10* - Гц мембраны все меньше и меньше перезаряжаются, в итоге емкостное сопротивление биологической ткани падает. Содержимое клеток все активнее включается в процесс образования ионных токов, т. е. проводимость ткани продолжает возрастать, а дюлектрическая проницаемость ее уменьша-12 . При этом значительно возрастают поляризация молекул и обусловленные ею токи смещения, что приводит к увеличению суммарных токов в тканях. При частотах ЭМ-колебаний вьпие 10" Гц емкостное сопротивление мембран клеток становится настолько малым, что клетку можно считать короткозамкнутой. Поляризация молекул и токи смещения становятся преобладающими. Характеристические частоты молекул воды, как связанной (гидратированной), так и свободной, а также относительно свободных белковых молекул крови оказываются равными или ниже частот воздействующего поля. Диапазон частот 3002000 МГц называют областью 5-дисперсии, обусловленной вращением боковых групп белковых молекул, а также релаксацией белковосвязанной воды [141]. За 7-дисперсию ответственны дипольные молекулы воды. Возбужденные молекулы приходят в колебательное движение, сталкиваются с псевдовозбужденными, передавая им свою энергию, которая расходу ется на химические преобразования, тепло и т. п. В итоге проводимость вновь резко возрастает, а дюлектрическая проницаемость меняется незначительно. На рис. 1.1 представлена также зависимость от частоты мнимой части комплексной диэлектрической проницаемости, которая понадобится в дальнейшем при оценке поглощения энергии биологическими объектами. Таким образом, с возрастанием частоты ЭМ-колебаний индуцирование ионных токов постепенно замещается поляризацией молекул. Например, диэлектрические потери в тканях при частоте 1 ГГц составляют примерно 50% общих потерь, а при частотах 10-30 ГГц - около 90 и 987с соответственно. В последние годы появились работы [32, 140, 141], юказывающие значимость клеточных мембран в биологических эффектах ЭМИ. Традиционные представления о мембране как изоляторе клеточного содержимого с удельной поверхностной емкостью порядка 0,1-3 мкФ/см и удельным поверхностным сопротивлением до 10* Ом/см при анализе ее микроструктуры претерпели некоторые изменения. При толщине отдельных мембран 50-100 А на каждой из них образуются разности потенциалов около 0,1 В (диффузионный потенциал Нернста), что составляет около 10* В/см. Кроме того, мембраны состоят из вытянутых цепных молекул (жидких кристаллов), которые в поле должны соответствующим образом перестраиваться. Все это приводит к нарушению действия привычного закона Ома в рассматриваемой области и может обусловить преимущественное поглощение ЭМ-энергии как на поверхности мембран (в основном вследствие потерь проводимости), так и в самой мембране (в результате электрических потерь). Однако такой механизм микрораспределения поглощенной энергии в биологической ткани не влияет на ее макроскопические диэлектрические свойства. Полная циэлектрическая проницаемость и удельная проводимость жаней заметно изменяются даже при небольших изменениях содержания воды, поскольку и диэлектрическая проницаемость и проводимость водь! значительно вьпие, чем у жировой или костной ткани. Диэлектрические свойства тканей зависят и от их температуры. СВЧ-диапазоне относительное изменение удельной проводимости и ди электрической проницаемости биологической ткани Да/а=2%/С Де/е= 0,5%/° С соответственно [141]. 1.2. ПРОНИКНОВЕНИЕ, ПОГЛОЩЕНИЕ, ОТРАЖЕНИЕ ЭМ-ЭНЕРГИИ В СЛОЖНОЙ БИОЛОГИЧЕСКОЙ СИСТЕМЕ Когда кривизна поверхности биологического объекта велика по сра нению с длиной волны, можно обратиться к исследованию характер стик распространения плоской волны в биологических тканях с плоски ми слоями. Зная толщину слоев и электрические свойства тканей, мож но легко рассчитать внутренние поля и токи, наводимые внешнил ЭМ-полем, коэффициенты отражения на границе между различными тка ними и распределение поглощенной энергии в различных слоях. Глубина d проникновения ЭМ-излучения в биологическую ткань обратно пропорциональная коэффициенту поглощения, определяет как расстояние, на котором амплитуда электрического поля убывае в е раз, а ППЭ - в раз [76,122, 140, 141], =Хо[2яе(71 + tg5- \)]"\ (1.1 где Хо - длина волны ЭМ-излучения в вакууме. При проникновении ЭМ-волны в ткань изменяется скорость ее рас пространения в среде, а следовательно, и длина волны в ткани Хтк роволны с частотой 2,4 ГГц могут проникать в ткань на глубину до 2 см, а микроволны с частотой вьппе Ю ГГц поглощаются в основном кожей человека. В тех случаях, когда толщина нескольких слоев вещества меньще глубины проникновения ЭМ-поля, часть энергии отражается от границ их раздела и в тканях возникают стоячие волны и связанные с ними максимумы поглощения. Анализ таких распределений подробно проведен в ряде работ по изучению и моделированию взаимодействия ЭМ-поля с биологическими объектами. Однако в реальных условиях поверхности раздела тканей имеют различные е, что затрудняет использование таких данных. Хтк = Хо [- (vl + tgS + l)] (1.2) Таким образом, на очень высоких частотах квадрат тангенса угла потерь значительно меньще единицы и Хтк X \fe следует из соотношения (1.2). При этом соотношение (1.1) показывает, что глубина проникновения ЭМ-излучения в биологическую ткань будет стремиться к нулю, приводя к поверхностному взаимодействию ЭМ-поля с биологи ческим объектом. Из-за высокой диэлектрической проницаемости ткани длина волны в ней оказывается меньше, чем в вакууме (рис. 1.2): в 6,5-8,5 раза для тканей с высоким содержанием воды и в 2-2.5 раза для тканей с низким содержанием воды [76, 141]. Ясно, что с повышением частоты глубина проникновения ЭМ-поля в ткани снижается. Так, мик- 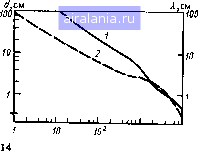 10 f, МГц Рис. 1.2. Изменение длины волны (7) глубины проникновения (2) ЭМ-волн в тканях с высоким содержанием воды в зависимости от частоты колебаний ЭМ-поля [140, 141) 1.3. ВЗАИМОДЕЙСТВИЕ ЭМ-ПОЛЯ С БИОЛОГИЧЕСКИМИ ОБЪЕКТАМИ (РОЛЬ ПОЛЯРИЗАЦИИ, РЕЗОНАНСА И ЗАЗЕМЛЕНИЯ В ОЦЕНКЕ ПОГЛОЩЕННОЙ ДОЗЫ) Общее количество поглощенной энергии и ее распределение внутри биологического объекта являются сложной функцией электрических свойств тканей, их общих геометрических размеров и условий облучения. Связь между падающей в единицу времени на единичную площадь тела энергией ЭМ-поля и возникающим при этом распределением удельного поглощения ЭМ-энергии не является постоянной. Дозиметрические исследования пространственных распределений удельного поглощения мощности (УПМ) необходимы для оценки поглощенной интегральной и локальной дозы, экстраполяции биологических эффектов с животных на человека. Распределение ЭМ-энергии в диэлектриках любой формы в общем случае описывается уравнениями Максвелла. Электрические свойства и геометрические размеры объекта в принципе позволяют рассчитать коэффициенты отражения на границах раздела различных тканей и воздуха, а также внутренние поля и наведенные токи в зависимости от характеристик внещнего ЭМ-поля. Уменьшение поля внутри плоского слоя вещества относительно прилагаемого внещнего поля определяется модулем комплексной диэлектрической проницаемости Ао/в = = evl + (tg5)- При ограниченных размерах объекта внутрейнее ЭМ-поле будет сложным образом зависеть от формы, размеров этого объекта и его ориентации относительно векторов поля. Как уже отмечалось, при низких по сравнению с высокими часатотах время релаксации свободных и связанных зарядов внутри биологической ткани мало по сравнению с периодом колебаний прилагаемого поля (е* велико), что приводит к значительному уменьшению поля внутри объекта. К настоящему времени появилось достаточно много интересных работ, посвященньк этой проблеме. Прежде всего следует отметить работы Шваиа, Гая, Дерни, Гандхи и их коллег [76, 100, 106, 107, 109, 111, 116, 120, 123, 140, 141], посвященные оценке роли поляризации, резонанс- 0 [1] 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 0.0094 |