
|
|
|
Главная -> Потоки электромагнитных излучений 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 [17] 18 19 20 21 22 23 24 25 26 27 28 ми воды выше проводимости чистой воды и электролитов. Этот эффекк иметь биологическое значение. Насыщение диэлектрической проницаемости белка происходит при 50 Поэтому изменения структуры протеина, наблюдаемые при напряженно уровня насыщения, по мнению Швана [76, 140), носят при нормальной темп спонтанный характер. Процессы релаксации, связанные с колебательными и крутильными : молекул, с нелинейностью диэлектрических свойств среды в условиях конк щего действия теплового движения молекул при достаточно высоких темпе приводят к возникновению обратимой поляризации. В полях напряженное ка нескольких вольт на сантиметр вряд ли могут появиться необратимые нз Результаты исследований приводят к важному выводу о том, что уде кость всех биологических мембран, в том числе клеточных и внутрикх органелл типа митохондрий, составляет 1 мкФ/см . Она практически не за частоты радиочастотного диапазона. В области низких акустических часто ная емкость мембран с уменьшением частоты возрастает. Можно оценить разность потенциалов на мембране, возникающую в СЕ При напряженности поля в живой ткани 1 В/м (СВЧ-излучение 10 мВт/с* удельном сопротивлении ткани примерно 100 Ом см плотность тока в ме составляет 10 мА/см. Наведенная разность потенциалов на мембране при/j равна 0,5 мкВ и с повышением частоты снижается. Это примерно в 1000 ] уровня потенциала, который принято считать биологически действующим ( Со Шваном не согласен Фрей [104], который считает, что биологическ ным значением можно считать напряженность поля менее 1 мВ. Для / </e1 разность потенциалов, приложенная к клетке, приходится на емкость ме Для клетки размером более 10 мкм в низкочастотном поле напряженностью! эта разность составляет несколько милливольт, что весьма существенно зрения биологического воздействия. Поскольку на низких частотах удельнов тнвление тканей лежит в пределах 500-1000 Ом-см, плотность токов равна 1 мА/см, а значение теплового шума на мембране - примерно 1 мкВ. Чтобы воздействовать на биополимеры, напряженность внешних эле» полей должна превышать напряженность силовых локальных полей, дос 1,5 кВ/см на расстоянии 100 А в многовалентных ноиах, и 1,8 кВ/см на расстоянии в молекулах гемоглобина. Разность потенциалов, возникающая на мембране в результате тиш1чных ловых воздействий СВЧ-полей, пренебрежимо мала и сопоставима с мемв шумом, который составляет примерно 1 мкВ. Метаболическая активность* ского мозга настолько высока, что его клетки излучают электромагнитную : 30 мВт/см. Только по одному радиочастотному диапазону ППЭ ЭМ-поля, излучается телом человека, составляет примерно 5 мкВт/см , Для прев уровня тепловых возмущений необходима слишком высокая напряженно Для описания иегепловых воздействий СВЧ-поля на биологические объек и Фостер [141] предлагают несколько возможных механизмов. Слабые роятнее всего, будут воздействовать на объекты, обладающие либо боль метрическими размерами, либо большими постоянными или индуцир дипольными моментами. Это возможно прн очень низких частотах. Большие размеры, необходимые для обеспечения реакции биологнче стемы на слабые СВЧ-поля, могут реализовываться в результате коопе взаимодействия нескольких клеток или макромолекул с импульсом СВЧ ния. Такое взаимодействие увеличивает эффективный размер структуры ветственно снижает порог возникновения рассматриваемого явления. По механизм кооперативного взаимодействия может быть обусловлен слабо св ми с поверхностью мембраны ионами и наличием в ней рыхлого каркаса из : ных полисахаридов. Возможно, что внутри мембраны протекают процесс нансного ЭМ-взанмодействия, которые лежат вблизи 100 ГГц. О биологи действии миллиметровых волн, в основе которых могут лежать резонансиь ния, сообщал Н. Д. Девятков [23]. Многие исследователи подчеркивают ело изучения биологического действия миллиметровых волн. Об этом, в ча ворил Партлоу на симпозиуме по биоэлектромагнетизму, который состоялся ""сиЭТле (США) в 1979 г. (112). Эти исследования в основном проводились на кле-очных суспензиях и культурах. Что же касается воздействия миллиметровых волн животных, то здесь вообще ничего не ясно, хотя биологические эффекты, очевид-могут возникнуть на уровне кожных реакций (при больших ППЭ), близких о характеру к реакциям, вызванным воздействием инфракрасного излучения. В последнее время появились высказывания о сверхпроводимости в органиче-материалах при комнатной температуре и выше. Ю. А. Холодов и М. А. Шиш-по [1 пь1тались использовать это явление для объяснения слабых (информациои-цух взаимодействий ЭМИ с биологическими структурами. В подтверждение не-специфического действия ЭМИ на животных они приводят данные Залуда и Воцеха гЦ], которые отмечали, что крысы выбирают камеру с СВЧ-облучением с ППЭ ниже 1 мВт/см , когда температура окружающей среды составляет 20 °С, и избе-fjjoi ее при 30°С. Во всех случаях повышения ректальной температуры ие наблюдалось. По мнению Барнса, взаимодействие ЭМ-излучения с биологической средой на уровне отдельной молекулы хорошо трактуется с позиций квантовой механики. Законы кинетики химических реакций применимы при изучении влияния ЭМ-излу-яений на движение молекул. Наиболее интересная область эффектов внешнего ЭМ-поля - это изучение состояний простейших структур, таких как мембраны, инфраструктуры клетки и граница раздела между цитоплазмой и мембраной. При описании эффектов ЭМ-излучения на молекулярном уровне определяющим является значение кванта СВЧ-излучеиия. Для доля частотой 10 ГГц оио составляет hv=l<r* Дж = 10" эВ. Переход кванта такого значения с одного уровня иа другой не может иметь биологического знане-ния. Значение ППЭ должно быть больше уровня теплового потока для того, чтобы в облучаемой системе могли произойти изменения. ППЭ должна составить /= = 10" Д/, где Д/«*1 МГц. Этот поток должен быть равным или большим 1 мкВт/см - нижний порог возможных биологических эффектов СВЧ-диапазона ЭМИ. Если действительно существуют "слабые" взаимодействия, ответственные за какие-то внешние эффекты, то вопрос заключается в том, каким образом и с помощью каких феноменов можно превысить тепловой шум биологической системы. Эйди [85] предлагает следующую концепцию. Под влиянием ЭМ-полей нарушаются связи ионов кальция с мозговой тканью. Это предположительно наблюдается на поверхности клеточных мембран. Почти на 2% увеличивается выход ионов кальция к медиатора ГАМК. При сверхнизких частотах его выход был наибольшим при частотах 6,16 Гц и напряженности поля 10 и 50 В/м соответственно или при модулировании частотой 9-16 Гц несущей частоты 147-450 МГц и интенсивности около 1 мВт/см. При интенсивности менее 1 мВт/см выход ионов кальция уменьшается. Дать биофизическое объяснение обнаруженным явлениям на основе существующих Представлений очень сложно. Например, поле частотой 16 Гц, напряженностью в воэ-пухе 50 В/м индуцирует в ткани поле напряженностью 10" В/см, а для создания 8 ткани напряженности 10" В/см (иапряжениость ЭЭГ) внешним полем с частотой 16 Гц необходима напряженность в воздухе 500 кВ/м. Главное место во взаимодействии биологической ткани с ЭМИ Эйди отводит •"•егочной мембране. Однако, чтобы слабые ЭМ-поля создавали в тканях напряженность намногоиже порога напряженности собственных электрических полей (Мембранный потенциал ю, синаптический потенциал 10 и ЭЭГ 0,1 В/см) и вызывали биологический эффект, необходимо по-иному взглянуть на саму мембрану ак на "жидкостную мозаичную модель" с большим количеством внутримембраных частиц в пределах двойного липидного слоя и трансмембранными связями (Внешнее пол - отрицательные заряды на поверхности мембраны - мбранные Ферменты-ядро). Чтобы избежать энергетического несогласования воздействую-т"" полей иа уровне теплового шума биологической ткани, Эйди предлагает рассматривать вначале эффекты взаимодействия ЭМИ с биологической тканью по по-ерхиости мембраны, а затем передачу информации внутрь клетки. Это осуществляется через такие ферменты, как аденилциклаза, которая заме- щает механизм превращения АТФ в АМФ и цАМФ. Все это в конечном счете з печивает запуск других ферментных систем маломощным воздействием ЭК чения. Между тем процессы, идущие затем внутри клетки, в 10-10* раз шают его по мощности. Передача энергетического сигнала с поверхности меь внутрь клетки осуществляется ионами кальция и соответствующей транс системой. Для объяснения слабых взаимодействий, по мнению Эйди, можно исг дисперсные, сопитонные модели. Явления солитонов, в частности А. А. Давь использованы при описании мышечного сокращения. Рассеяние солитонов в i лах при воздействии ЭМИ можно объяснить следующим образом. Эпектрок ное излучение деформирует электронные орбитали атомов, смещает ядра, ко совершают нелинейные колебания, волна зтих колебаний распространяется молекулы со скоростью звука. Двигаясь таким образом, солитон отдает в виде излучения типа Черепковского. Солитон может быть рассеян на не ностях и направлен вдоль ответвлений молекулы. Энергия, доставленная сол на удаленный конец молекулы, может вызвать вторичные химические Применительно к ЭМИ можно предположить, что электромагнитная волна j ся, опережая волну изменений в липидных молекулах вдоль мембраны, и вь вторичные процессы. Это пример того, как начальное событие, локализованное! мембраны, инициирует трансмембранные процессы с последующими вто реакциями внутри мембраны. Мы специально остановились на рассмотрении механизмов слабых вза ВИЙ, поскольку на их основе делаются попытки объяснить многие феноме воздействии ЭМ-излучения на ЦНС. Можно справедливо задать вопрос: модул ные или низкочастотные ЭМИ оказывают на организм вредное воздействие i имеют для организма прежде всего физиологическое значение. Скорее всего i нее более справедливо. Факты, полученные такими крупными нейрофизно как Эйди и др., нельзя игнорировать, но они пока что имеют лишь научный i Более того, они трудновоспроизводимы. Эти феномены, возможно, и наб в слишком узких областях модуляций, но они требуют очень корректной i Наиболее четкие изменения в организме возникают тогда, когда во ЭМИ сопровождается тепловыми эффектами. Последние, с нашей точки должны быть ведущими при оценке профессиональной вредности радиоча излучений. 5.2. ТЕПЛОВОЙ СТРЕСС ПРИ МИКРОВОЛНОВОМ ОБЛУЧЕНИИ Совершенно определенно можно сказать, что хорошо изучек стоверным фактом является тепловой эффект ЭМИ, особенно мк нового диапазона [4, 46, 141, 126, 154]. На основе литературных , можно заключить, что с увеличением интенсивности и времени 3M5J чения у мьпней, крыс и собак повышается температура тела, ется частота сердечных сокращений и дыхательных движений. В ус многократного облучения наблюдается, по-видимому, эффект те адаптации, при небольших интервалах между воздействиями во функциональная кумуляция. Изучение теплового стресса при воздействии ЭМИ целесообр чать с рассмотрения основны.-с физиологических сдвигов при ППЭ, т. е. теплового баланса организма, определяемого по ре температуре, локальной температуре регулирующих центров, сердечно-сосудистой и дыхательной систем, установления кс между тепловым балансом и уровнем смертности животных и ог ния порогового значения подъема температуры при вероятности 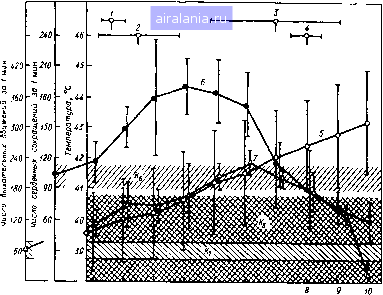 4 5 в 7 Время облучения, мин Рис. 5,1. Время возникновения клинических симптомов и некоторых физиологических сдвигов в период микроволнового облучения (300 мВт/см ): 1 ~ повизгивание; 2 ~ двигательное возбуждение; 3 - выраженный визг; 4 -судороги; 5 - температура тела; 6 - пульс; 7 ~ дыхательные движения; К-К-, -контрольные значения для 5- 7 соответственно 0,1%, а также с рассмотрения эффектов восстановления и кумуляции при тепловом воздействии ЭМИ. При общем облучении хорошо прослеживается последовательность изменений основных физиологических параметров и поведения собаки (рис. 5.1). Наибольшая корреляция наблюдается между изменением ректальной температуры и частотой дыхательных движений. Это следует и из сопоставления данных по изменению частоты дыхательных движений, частоты сердечных сокращений при ППЭ, равной 100, 300 и 500 мВт/см (рис. 5.2). При анализе экспериментальных данных установлена корреляция между вероятностью гибели собак и градиентом нарастания ректальной температуры (ДГ), которая в то же время не зависела от ППЭ (рис. 5.3). При вероятности гибели 99,9% Д7" составил 4,78° С, при вероятности гибели 50 и 0,1% - 3,52 и 2,1б°С соответственно. Последнее значение прироста температуры особенно интересно тем, что оно практически соответ-•iByei пределам физиологических колебаний этого показателя у живот-Hbix. Рассмотрим такую аналогию. К. П. Иванов [28] вычислил прирост Температуры за счет собственного метаболизл4а и при отсутствии теплоотдачи в околоклеточную среду. По его Данным, 1 г вещества тел ней-Ронов выделяет примерно 1,8 к ал/(г мин) тепла. Тогда Д 7- = Q/Mc = 1,8/0,83 = 2,16° С, где Q - тепло, кал/(г мин); М - масса, г; с - теплоемкость, 0,83 кал/(г "С). Однако тепло от тел нейронов (в слзае, если ступления тепловой знергии извне) отводится очень быстро и Д 7" i ляег только 0,6-10"°С [28]. Если проанализировать зкспериментальные данные по изме ректальной температуры в период облучения у собак (рис. 5.4) в установления корреляции между интенсивностью и временем облу il 1
40- SO .,200 J М J I,  I I I I I I I I I I M I I I I I I I I I I I I I , .......I « 20 SO 40 Время облучения, мин 60 70 Рис. 5.2. рульс и дыхание (по отношению к максимальному значению 500 мВт/см ) у собак в период микроволнового облучения; 1-3 - ППЭ 500, 300, 100 мВт/см соответственно. Заштрихованные соответствуют 0,1-99,99ной гибели животных; К - контрольные значения  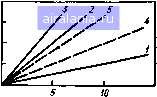 S00 , да, мВт/см Рис. 5.3. Прирост ректальной температуры (ДГ) и гибель собак в зависимости от ППЭ микроволнового облучения Рис. 5.4. Прирост ректальной темпе при микроволновом облучении собак:1 1-3 - ППЭ, равная 100, 500 мВт/см соотвегственно (соб ные данные для 2,4 ГГц); 4 и 5 - равная 220 и 330 мВт/см для 0,2 [861 3 5, Зависимость ППЭ от времени t г*" роволнового облучения собак для ГгГи " 2 " "Р" постоян-н приросте температуры ДГ, равном 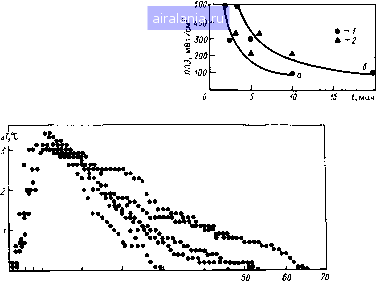 0 24 6 8 10 20 30 40 Время о6л1/чения,мш1 Время после облучения, мин Ркс. 5.6. Прирост и спад ректальной температуры ДГу крыс (масса 240 г) во время об/ц-чения и после него с ППЭ 100 мВт/см (2,4 ГГц) ИЛИ дозой при определенном значении ДГ, то получим практически параболическую зависимость (рис. 5.5) /Г = 50 Дж/см (для Д7=1С) и /f=105 Дж/см (для ДГ = 2°С), где / - ППЭ, мВт/см, и t - время облучения, с. Минимальный порог интенсивности, при котором справедливость зтих [соотношений может сохраняться, не должен быть ниже 80 мВт/см (см. .5.3). После окончания облучения спад ректальной температуры идет зкспо-(енциально. Отмечаются довольно большие индивидуальные колебания ремени восстановления (рис. 5.6). Естест-•енно, что, как и всякая система с обратной связью, теплорегуляция не является исклю-ением. После достижения исходного уров- температура, как правило, падает ниже контрольных значений. Это показано на мы-крысах и собаках. Любую температур-гУю кривую при действии ЭМИ можно раз- Иить на четыре участка: подъем температуры г° Время облучения, дальнейший небольшой 1 Продолжительность плато (7) и величина ,Р°«а ректальной температуры у собак (2) Исимости от ППЭ 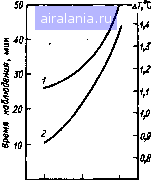 О 100 т 500 Плотность потока энергии, мВт/см 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 [17] 18 19 20 21 22 23 24 25 26 27 28 0.01 |